férencient en neuroblastes et cellules
gliales dans la région périventriculaire
(également appelée zone germinative
périventriculaire ou matrice germina-
tive). A la suite de la dernière division
mitotique, les neurones corticaux
migrent à distance de leur région d'ori-
gine, selon 2 modes de migration: la
migration radiaire, qui intéresse princi-
palement les futurs neurones néocorti-
caux, et la migration tangentielle, qui
concerne les interneurones GABAer-
giques. La majorité des neurones migrent
à partir de la région périventriculaire, de
façon radiaire selon les fibres de glie ra-
diaire, vers la surface piale. En arrivant
au niveau de cette surface piale, les neu-
rones s'organisent en couches succes-
sives, selon un processus de lamination
extrêmement complexe. Ces vagues mi-
gratoires des neurones néocorticaux s'ef-
fectuent entre la 20
ment qualifié de «inside-out», qui per-
mettra la formation du cortex mature en
6 couches. Dans un premier temps, les
neurones les plus jeunes forment une
«préplaque», qui correspond à une orga-
nisation précoce des neurones à la sur-
face. La plaque corticale «définitive» se
formera secondairement, à l'intérieur de
cette «préplaque»; ainsi, les neurones
nouvellement formés divisent la pré-
plaque en 2 parties: la zone marginale
externe et la sous-plaque. Par la suite, les
neurones qui migrent en premier s'ar-
rêtent dans les couches les plus pro-
fondes. Les neurones suivants traversent
la sous-plaque, les neurones «arrivés
avant eux», et se placent en superficie
pour former les couches corticales plus
superficielles, de façon adjacente à la
zone marginale, et ainsi de suite pour les
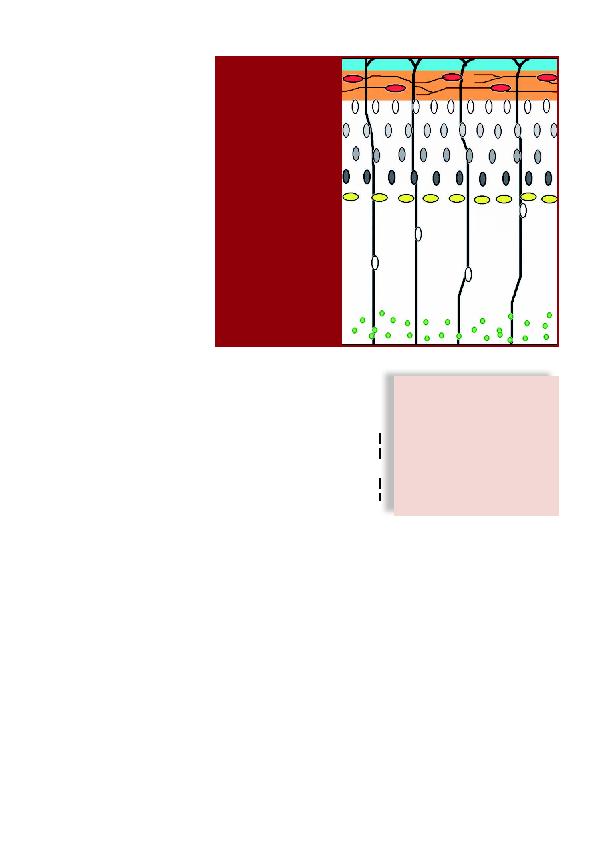
vagues de migration suivantes (Figure 1).
En revanche, la plupart des neurones qui
formeront les interneurones GABAer-
giques migrent de façon tangentielle,
gner leur localisation appropriée du cor-
tex; par exemple, la migration tangen-
tielle des cellules de Cajal-Retzius à par-
tir de l'éminence ganglionnaire vers le
cortex, ou le courant migratoire rostral
qui interconnecte la région périventricu-
laire aux bulbes olfactifs. Une fois que
les neurones arrivent à leur destination
finale, ils arrêtent leur migration et s'or-
ganisent selon des patterns architecto-
niques spécifiques qui obéissent à des
voies de signalisation complexes, gui-
dant les neurones vers leur localisation
finale dans la couche corticale. La 3
tion finale en 6 couches successives, as-
sociée à la synaptogenèse, avec une pro-
lifération des neurites, et des processus
de stabilisation comprenant la mort neu-
ronale programmée, la régression des
neurites et des synapses redondantes.
l'origine d'un dysfonctionnement corti-
cal majeur, qui résulte en un retard men-
tal parfois sévère et une épilepsie sou-
rité de ces encéphalopathies est liée au
type de malformation corticale, à son
étendue et à la présence d'anomalies
associées de la fosse postérieure et du
corps calleux. Leur diagnostic repose sur
une imagerie de bonne qualité, explo-
rant non seulement l'aspect du cortex et
la localisation maximale des anomalies,
mais également la présence d'anomalies
associées (substance blanche, ganglions
de la base, tronc cérébral et cervelet) qui
guide le diagnostic génétique et molécu-
laire. Selon le schéma de classification
proposé par Barkovich en 2005, on les
subdivise en fonction du mécanisme
neurobiologique principalement affecté
(Tableau 1). D'un point de vue physiopa-
inside-out durant la corticoge-
nèse: les neurones les plus
jeunes migrent et passent à tra-
vers les plus anciennement or-
ganisés, en utilisant la glie ra-
diaire comme guide (en vert,
les neurones de la région péri-
ventriculaire; en rouge, les cel-
lules de Cajal-Retzius, qui
constituent les cellules pion-
nières de la préplaque).