ba¤lanmas> ile iinhibitör alt üniteler bie dimer olarak iki katalitik alt
üniteden ayr>fl>r. Böylece katalitik üniteler ATP'nin gama pozisyo-
nundaki fosfat> proteinlerin serin ve threonin bileflenlerine tafl>nmas>-
n> katalize eder. Fosforilasyon arac>l>kl> bu kovalent modifikasyon bir
allosterik konformasyon dönüflümü ile substrat proteinin aktivitesini
de¤ifltirir. Hormon sinyali böylece enzim aktivite de¤iflikli¤i üzerin-
den hücre fonksiyonlar>n> düzenlemifl olur. Stoplazmik proteinlerin
fosforilasyonu glikoliz gibi de¤iflikliklere yol açar; aktifleflmifl katali-
tik kinazlar nükleusa göçerek transkripsiyon faktörlerinin fosforilas-
yon ve aktivasyonunu tetikler, örnek cAMP yan>t ünitesi ba¤lay>c>
proteini (CREB).
yonlar> geri dönüflümlü k>l>n>r. Fosfodiyestarazlar regülasyona tabidir
ve cAMP y>k>m>n> engelleyerek etkisini uzatan metilk>santin gibi inhi-
bitör ilaçlar>n hedef molekülüdür. Fosfotazlar fosforilasyon arac>l>¤la
regüle edilen fosfotaz inhibitör proteinleri taraf>ndan kontrol edilirler.
Memelilerde en iyi tan>mlanm>fl enzim atriyal natriüretik hormon re-
septörüdür (ANH). ANH ba¤lanma bölgesi reseptörünün ekstraselü-
ler k>sm>nda yerlefliktir, stoplazmik guanilat siklazdan tekli membra-
n> geçen bir bileflen ile ayr>l>r (fiekil 234-2). Adenilat siklaz>n aksine re-
septör ve katalitik etkiler ayn> molekülde yerleflmifltir. Aktivite birin-
cil olarak ligand ba¤lanmas> ile regüle edilir fakat ayn> zamanda enzi-
min fosforilasyonuna da ba¤l>d>r, defosforilasyon ile duyarl>l>k azal>r.
Guanilat siklaz>n stoplazmik bir formun hem grubu içerir ve nitröz ok-
sit ile serbest radikaller taraf>ndan aktiflefltirilir. cGMP, cGMP ba¤>m-
l> protein kinazlar>n kontrol bölgelerine ba¤lanarak etki gösteririr. G-
kinaz, bir dimerik enzim olup A-kinaz ile evrimsel olarak iliflkilidir,
cGMP ba¤lanmas>yla allosterik biçimde aktifleflir. A-kinaz gibi enzim
fonksiyonlar>n> de¤ifltiren ve fizyolojik cevaplar oluflturan protein fos-
forilasyonlar>n> katalizler. Reaksiyonlar cGMP fosfodiyestaraz ve pro-
tein fosfotazlarca sonland>r>l>r. cGMP fosfodiyestaraz kalsiyum-kal-
modulin ba¤lanmas>yla aktifleflir.
ve diaçilgliserol (DAG). PI döngüsü bu fosfolipidin yap>m, y>k>m, ye-
niden sentezini kapsar. PI üç karbonlu gliserol iskeleti ile birinci ve
bondaki fosfoester ba¤>ndan esterifiye inositol halkas>ndan oluflur.
Farkl> kinaz enzimleri inositol halkas>n>n üç, dört, beflinci pozisyonla-
r>n>n fosforilasyonunu katalize eder. Temel fosforilasyonlar s>ras>yla
önce dört sonra beflinci pozisyonda gerçekleflir. Aktifleflmifl hormon
reseptörlerinin fonksiyonu fosfoinositidaz (fosfolipaz C) enzimini
uyararak fosforlanm>fl inostiol sal>n>m> sa¤lamak ve inositol trifosfat
(IP3) ve DAG oluflturmakt>r. IP3 stoplazmadaki kalsiyum konsantras-
yonunu art>r>r.
yumu serbestlefltirir. Bazal stoplazmik kalsiyum konsantrasyonu hüc-
re d>fl> ve depo bölgelerinden yaklafl>k bin kat daha düflüktür. Depola-
r>n boflalmas> ve hücre içine kalsiyum girifliyle stoplazmik kalsiyum
h>zla artar.
masyonunu de¤ifltirerek kalsiyum-kalmadulin kompleksinin çeflitli
enzimlere ba¤lamas>n> uyar>r. Kalsiyum-kalmadulin düz kas kon-
traksiyonlar>nda rol alan myozin hafif zincir kinaz>; glükojen y>k>-
m>nda rol alan fosforilaz kinaz; sinaptik iletide rol alan kalmodulin
ba¤>ml> protein kinaz gibi baz> protein kinaz enzimlerini kontrol
eder. Kalsiyum-kalmadulin cAMP ve cGMP konsantrasyonlar>n> et-
kileyen siklik nükleotit fosfodiyestaraz, adenilat/guanilat siklaz en-
zimlerini regüle eder ve mikrotübül birleflmesinde rol al>r. Dolay>-
s>yla kalsiyum-kalmodulin stoplazmik kalsiyum konsantrasyonu
üzerinden tafl>nan bilgilerle çok çeflitli proteinlere ba¤lanarak onla-
r>n etkilerini kontrol eder.
yaç duyar, böylece bu yola¤>n ikincil mesajc>lar> enzimin aktivitesini
art>rmak için birlikte çal>fl>rlar. Aktif forbol esterleri gibi tümör uya-
r>c>lar birer DAG analo¤u olup protein kinaz C üzerinden etki göste-
ririler.
foinositidaz enzimi G proteinlerce aktifleflirken bir di¤eri trozinkinaz
ba¤lanmas>ylfla aktifleflir ve tirozin fosforilasyonuna maruz kal>r. Di-
¤er kinazlar inositol halkas>n>n farkl> pozisyonlar>n> fosoforlar; PI 3-
kinaz baz> özel tirozinkinazlarca aktiflefltirilerek kalsiyum mobilizas-
yonundan farkl> fonksiyonlar>na sahip PI metaboliklerini aç>¤a ç>ka-
r>r. 4,5 pozisyonlar>ndan fosforlanm>fl PtdIns, PH bölgelerini tan>-
mak, kinazlar ve di¤er proteinleri lokalize etmek aktiflefltirmek fonk-
siyonlar>n> üstlenmifltir, ve PtdIns3P, proteinlerin FYVE bölgelerine
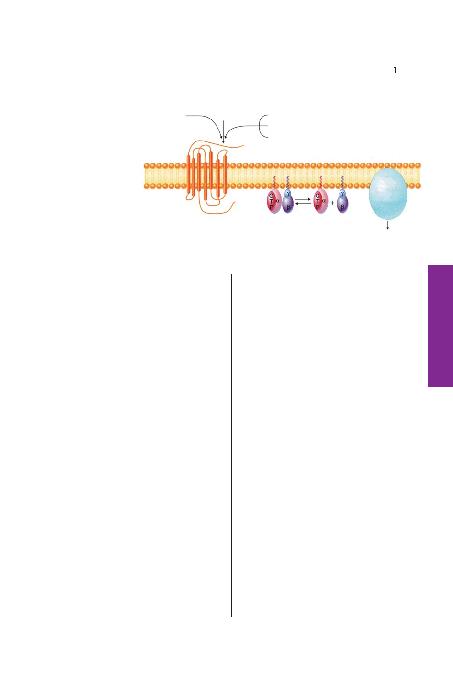
7'li membran reseptörlerine ba¤lana-
rak G-proteini aktiflefltirip; GDP-GTP
de¤iflimini tetikler. GTP ba¤lam>fl G-
protein farkl> yolaklar üzerinden sinyal
iletir. GTP'nin hidrolizi ile bu mekaniz-
ma sonlan>r. (Bockaert J, Pin JP: Mol-
lecular tinkering of G protein-coupled
receptors: An evolutionary process.
EMBO J 1999;18: 1723-1729'dan al>n-
m>flt>r).
· ACTH, TSH,
· Sitokinler
· Biyojenik aminler
· Prostaglandinler
· Nükleotidler/
· Adenilat