kor bu gen parçalar>n>n farkl> kombinasyonunu kullan>r.
ya da variable/ba¤lay>c> birleflimlerinde nükleotid de¤iflikliklerine ne-
den olurlar. Bu kusurlar nedeniyle tamamen farkl> amino asit dizileri
ile sonuçlanabilirler. Son olarak, deoksiribonükleotidil transferaz enzi-
mi taraf>ndan birleflme bölgesine rastgele nükleotidler eklenebilir.
tör redaksiyonu), immün yan>t s>ras>nda variable/diversity/ba¤lay>c>
ünitelerin farkl> sabit bölge genleri ile kombine oldu¤u s>n>f de¤iflim-
leri, ya da immün yan>t s>ras>nda tümleyici-belirleyici bölgelerin mu-
tasyona u¤rad>¤> somatik hipermutasyonlar> içerebilir.
s>n>f>, iki farkl> T hücre alt grubu taraf>ndan tan>n>r. CD4
nel olarak, MHC s>n>fII moleküller, yaln>zca dendritik hücreler, mono-
sitler, makrofajlar ve B hücreler gibi profesyonel antijen sunan hücre-
ler üzerinde eksprese olurlar. MHC s>n>f II molekülleri ile sunulan
peptidler, hücre d>fl> antijenik yap>lar olup önce endozomlar içine al>-
n>rlar. Endozomlarda özellikle katepsin olmak üzere proteinazlar tara-
f>ndan sindirilerek sunulmak üzere haz>rlan>rlar. Bazen hücre içi pro-
teinler veya membran proteinleri de bu yolla sunulabilirler. MHC s>n>f
II moleküller invaryant zincir ad> verilen bir protein ile iliflkili olarak
endoplazmik retikulumda toplan>rlar (fiekil 42-3). Moleküller, peptid-
ba¤lay>c> yar>¤>n hücre d>fl> proteinlerden türeyen peptidlere ulaflabi-
lir hale gelmesi için invaryant zincirin yar>ktan ç>kar>ld>¤> endozoma
lekülleri, CD4
lar (fiekil 42-3). Antijen iflleyici proteinler ile iliflkili tafl>y>c> olarak ad-
land>r>lan, özelleflmifl bir tafl>y>c> sistem, antijenik peptidlerin sitozol-
den endoplazmik retikuluma geçiflini kolaylaflt>nr. Burada yeni olufl-
turulan MHC s>n>f I moleküllerine ba¤lanan peptidler, antijen-spesifik
CD8
CD4
antijenler, y>k>larak CD8
hücre olgunlaflmas>n>n ileriki basamaklar>n>n gerçekleflece¤i timusa yer-
leflmifllerdir (fiekil 42-4). T hücreler, TCR genlerini rekombine edebilme-
lerini sa¤layan rekombinaz aktive edici genler ve terminal deoksinükle-
otidil transferaz eksprese ederler. TCR'nin yeniden düzenlenen
lat>r. Bunu takiben, TCR
çevre, özellikle de timik epitelyal hücreler, makrofajlar ve dendritik hüc-
reler, taraf>ndan ayarlanan bir çok farkl>laflma ve seçilme basamaklar>n-
dan geçerler. Timositlerin erken evreleri, ço¤unlukla epitel hücrelerle bir-
birlerini etkiledikleri timik kortekste kal>rlar. Sonras>nda, kortikomedül-
ler bileflkede dendritik hücreler ve makrofajlarla karfl>laflt>klar> medulla-
ya göç ederler. Timik stromal hücreler IL-7 gibi lenfopoetik büyüme fak-
törleri salg>layarak T-hücre proliferasyonunu düzenlerler. TCR'nün epi-
telyal hücreler ve dendritik hücreler/makrofajlar üzerindeki MHC mo-
lekülleri ile birbirini etkilemesi timositin gelece¤ini belirler. Timik epitel-
yal hücreler üzerindeki peptid/MHC komplekslerinin TCR taraf>ndan
düflük düzeyde tan>nmas> "pozitif seleksiyon" ile sonuçlan>r. Bu tan>ma
olay>, hücreleri apoptotik hücre ölümünden kurtar>r ve sadece fonksiyo-
nel reseptörleri olan T hücrelerin yaflamas>n> sa¤lar. Hiçbir MHC antijen
kompleksine uymayan reseptör tafl>yan timositler ise ihmal edilerek ölür.
TCR ve peptid/MHC kompleksi aras>nda yüksek afiniteli iliflki, bunu
gösteren T hücrenin apoptotik ölümüne neden olur. Bu "negatif seçilme"
olay>, kendi-antijenlerine özgüllük gösteren T hücrelerini yok eder ve bir
çok otoantijene karfl> "santral tolerans"tan sorumludur. Timositlerin yak-
lafl>k %1'nin bu s>k> seçilme olay>n> geçebildi¤i tahmin edilmektedir. Se-
çilme geçerken, T hücreleri, hücre yüzey moleküllerinin s>ral> ekspreyo-
nu ile farkl>laflmay> sürdürmektedir. CD4 ve CD8 moleküllerini ekspre-
se eden timositler, MHC s>n>f II kompleksler ile iliflki kurmufl olan CD4
CD8
bafllar. T hücreler taraf>ndan antijenin tan>nmas> proliferasyon ve fark-
l>laflmay> sa¤lar ve farkl> efektör fonksiyonlar> tetikler. Ancak burada
yaln>zca TCR uyar>lmas> yeterli de¤ildir. T hücre yüzeyindeki aksesu-
var moleküllerin de antijen sunan hücre üzerindeki ligandlar> ile ilifl-
kiye girmesi gerekir. Aksesuvar moleküllerin bir spektrumu bilinmek-
tedir (Tablo 42-1). Koreseptörler, CD4 ve CD8, MHC s>n>f I ve s>n>f II
mokülleri ile iliflkiye girer ve TCR üzerinden aktivasyon sinyalle des-
tekler. Adezyon molekülleri (integrinler) T hücreler ve antijen sunan
hücreler aras>ndaki iliflkiyi stabilize eder. Son olarak, özelleflmifl ko-sti-
mulatör moleküller TCR sinyaline ek olarak ikinci bir sinyal daha sa¤-
lar. Bu ikinci sinyalin yoklu¤unda, T hücreler apopitozise u¤rar ya da
yan>ts>z yani anerjik hale gelirler.
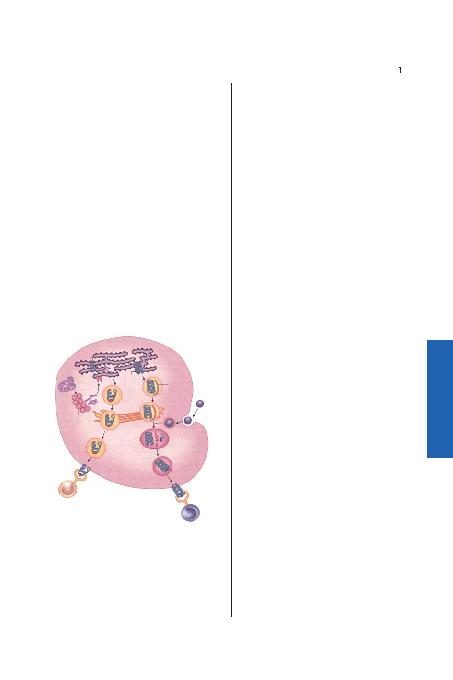
inler özelleflmifl peptid tafl>y>c>lar> (TAP) taraf>ndan endoplasmik retikulumda dö-
nüfltürülmüfl olan peptid fragmanlar>n> üretmek üzere proteozomlar taraf>ndan
parçalanm>flt>r. Peptidler MHC s>n>f I moleküllerine ba¤land>ktan sonra endoplas-
mik retikulumdan ayr>l>rlar ve golgi aparat>yla hücre yüzeyine do¤ru ilerlerler.
MHC s>n>f I/peptid kompleksleri CD8
veziküllere al>n>rlar. Endozomlardaki pH giderek azal>r antijenleri sindirecek olan
proteazlar aktive olur ve antijenler peptid fragmanlar>na dönüfltürürler. MHC s>n>f
II moleküllerini içeren veziküllerle birleflme sonras>nda antijenik peptidler antijen
ba¤layan olu¤a yerleflirler. MHC s>n>f II/peptid kompleksi CD4
zincir