1371.Circulating lipopolysaccharide, presumably from translocated intestinal flora,
is increased in patients with HIV
meta-analysis. Clin Infect Dis 2005; 41:713-720.This systematic review of eight tri-
als involving more than 6000 patients found that HIV+ patients coinfected with
HCV have a less robust CD4 response to potent anti-retroviral therapy
roviral therapy. Clin Infect Dis 2006; 42:1639-1646.Patients with more advanced
illness are at increased risk of developing immune reconstitution syndrome
re was no decline in HIV-1specific CD4 cells when ART was discontinued and
HIV viremia rose
Daha önceden keflfedilen insan ve hayvan retrovirüslar>ndan (Bölüm
401) köken alan HIV'>n bulunmas> büyük bir bilimsel baflar> idi. Has-
tal>kl> kiflilerde CD4
gibi, AIDS'li hastalar>n doku kültürlerinde etiyolojik ajan HIV-1'in
varl>¤> ve T lenfositlerin y>k>m> gösterildi.
nin tersine HIV-1 sinsitya oluflumuna ve hücre ölümüne sebep olarak
lenfositler için belirgin sitopatik özellikler gösterir. Morfolojik olarak
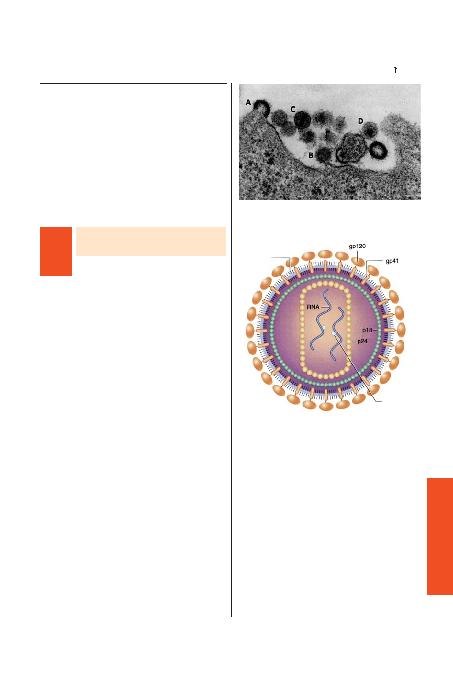
HIV-1, HTLV-1 ve di¤er onkojenik retrovirüslerden lipid bir zar etra-
f>n> saran yo¤un, silindirik kor varl>¤> ile farkl>l>k gösterir (fiekil 409-1).
RNA ba¤>ml> DNA polimeraz ya da revers transkriptaz virion koru-
nun içinde paketlenmifltir ve tek sarmall> RNA genomunu, konak hüc-
re genomuna entegrasyon için öncül molekül olarak görev yapan çift
sarmall> DNA'ya dönüfltürür. HIV-1'in ana yap>sal proteini p24 kap-
sid proteini ve p18 matriks proteinidir. Viral kor protein yap>lar>n> et-
raf>nda, hücre yüzeyinden viral tomurcuk olarak ç>kan konak d>fl za-
r>ndan kaynaklanan çift tabakal> lipid zarf bulunur. Virüs spesifik gen-
ler taraf>ndan kodlanan hücre ba¤lanmas>ndan ve giriflinden sorumlu
olan gp120 ve gp41 olarak isimlendirilen zarf glikoproteinleri bu d>fl
viral membran>n üzerinde bulunurlar.
l>d>r.
etkileflimin spesifisitesi viral gp120 zarf glikoproteinin hedef hücre ile
iliflkili CD4 molekülü aras>ndaki yüksek afiniteli etkileflim sonucudur,
bu etkileflim gp120'de konformasyonel bir de¤ifliklik ve sonras>nda
hücresel kemokin reseptörü ailesinin bir üyesi (CCR5 ya da CXCR4) ile
etkileflim oluflturur. Gp120, CD4 ve CCR5/CXCR4 aras>ndaki bu spesi-
fik etkileflimler viral ve hücresel zarlar>n birleflmesi ve nükleoprotein-
virüs kompleksinin içeri al>nmas> ile sonuçlan>r. Revers transkriptaz ta-
raf>ndan katalize edilen ters transkripsiyon nükleoprotein kompleksi
içinde viral RNA'n>n çift sarmall> DNA kopyas>n> oluflturur, bu DNA
nükleusa gider, nükleusta provirüs oluflumu için viral DNA konak kro-
mozomuna kovalent olarak ba¤lan>r. Daha sonra viral DNA ekspresyo-
nu, viral proteinler; viral DNA ve RNA düzenleyici elementlerle etkile-
flen kona¤a ait hücresel proteinler taraf>ndan kontrol edilir. Transkrip-
siyona u¤ram>fl viral RNA viral proteinlere çevrilir. Yeni virionlar ge-
nomik uzunluktaki viral RNA'n>n, revers transkriptaz>n, yap>sal ve dü-
zenleyici proteinlerin ve zarf glikoproteinlerinin topland>¤> yer olan
hücre yüzeyinde toplan>rlar. HIV-1 provirüs konak hücre genomuna
kovalent olarak ba¤land>¤>ndan provirüs konak genomunun sabit bir
komponentini temsil eder. Entegre provirüs transkripsiyonel olarak la-
tent kalabilir ya da patlay>c> tarzda bir progen virüs üretimi için yüksek
seviyelerde gen ekspresyonu yapabilir.
morfogenezin tüm aflamalar>nda gösterilmektedir; erken (A), geç (B) tomurcuk-
lanan formlar ve serbest matür virionlar (C ve D) santral kor ile birleflmifl olarak
görülüyor. Virion çap> yaklafl>k 110 nm'dir.
George V. Kelvin. 'dan al>nm>flt>r).
transkriptaz