PtdIns kinazlar böylece membranda PH, FYVE ve PX birleflimlerin-
den oluflan sinyal iletim kompleksleri için iskele bölgeleri oluflturur.
Glikosifingolipid metabolizmas>n>n bir bilefleni olan sifingozin pro-
tein kinaz C'yi inhibe eder ve bu proteinin ikili kontrolünü sa¤lar.
Özel fosfotazlar inositol halkas>ndan fosfat gruplar>n> kopartarak ak-
tivitesini sonland>r>r; lityum bu fosfatazlardan birisinin aktivitesini
bloke ederek biyolojik olarak aktif inositol fosfat birikimini uyar>r.
Kanserlerde s>k rastlanan PtdIns3 fosfataz geni (PTEN) delesyonlar>
"kapatma" kodonlar>n> ortadan kald>rarak büyüme sinyallerini aç>k
b>rak>r.
kil 240-2).
reseptör ve JAK2 ikinci s>n>fa aittir.
ba¤lan>r. Oysa pek çok kinaz hücre proliferasyonu, geliflim ve diferan-
siyasyon fonksiyonlar> tafl>yan önemli proteinlerin tirozin bileflenleri-
ni kovalent olarak modifiye eder. Bu grup reseptörlerin sistein'den
zengin ekstrasüleüler ligand ba¤layan bölgeleri ba¤lanma bölgelerin-
de monomer (EGF reseptörleri) veya dimerler (insülün reseptörü)
oluflturur yada immune globulin benzeri yap>lara dönüflür (PDGF ve
FGF reseptörler). Stoplazmik protein trozinkinaz bölgeleri ATP ve
substrat ba¤layan bölgeler içeren yüksek homoloji gösterir fakat farkl>
reseptörler farkl> substratlar> tan>r. Örne¤in insulin glikoz al>m>n> uya-
r>rken EGF hücre proliferasyonunu tetikler.
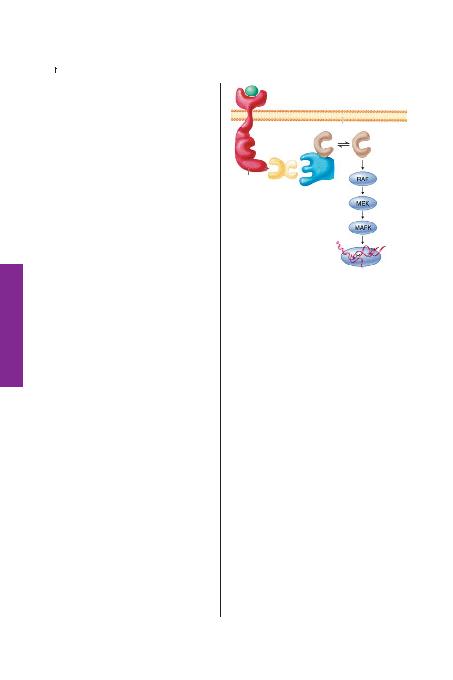
protein ras ile biten bir dizi sinyal iletim yola¤>ndan aktar>l>r (fiekil
240-4). Ligand ba¤lanmas>na cevaben reseptör trozinkinazlar ya ken-
dilerini fosforlar yada ba¤lay>c> substurat> fosforlar. 100 aminoasit
içeren, src, SH2'deki bir bölgenin homolo¤u olan proteinler tirozin
fosforilasyonun bu bölgelerine s>k>ca ba¤lan>r. Büyüme faktör resep-
tör ba¤layan protein 2 (GrB2) SH2 bölgesi içeren tirozin fosforilasyon
bölgesine tutunan bir arac> moleküldür. Shc bir baflka s>k kullan>lan
arac> moleküldür. Grb2 ayn> zamanda prolinden zengin guanin nük-
leotit de¤iflim proteini SOS'i ba¤layan iki adet sH3 bölgesi içerir. Bu
yüksek afiniteli protein-protein etkileflimi SOS'i, ras'>n inaktif olarak
yerleflti¤i hücre mebran>na tafl>r.aktifleflmifl GTP'ye ba¤l> ras daha
sonra bir raf-1 içeren serin treonin protein kinaz kaskad>na eklenir,
ard>ndan MEK ve MAP (mitojen aktifleflmifl protein) kinazlara tutu-
nur. Böylece bilgi gen ekspresyonu ve hücre bölünmesini kontrol et-
mek üzere domino tafl> etkisi yaratarak hücre içine yay>l>r. Reseptör
tirozinkinazlar ayr>ca SH2 bölgeleri içeren ilave sinyal yolaklar> ve
fosfolipaz C-gama ve transkripsiyonun sinyal ileticileri ve aktivatör-
leri (stat) ve PI3 kinaz içeren proteinlerin tirozin fosforilasyonununa
kat>l>r.
lasyonu (2) tirozin alt ünitelerinden fosfat koparan tirozin fosfotazlar
(3) tirozin bileflenlerinden ADP'ye fosfat tafl>yan ters kinaz reaksiyonu
(4) ras'a ba¤l> GTP'nin GDP'ye hidrolizi ligand ile aktifleflmifl tirozin-
kinazlar>n geri dönebilirli¤i ve regülasyonu önemlidir.
mesini sa¤layacak flekilde kontrol yolaklar>n> bypass edebilir. Ki-
nazlar gen amlifikasyonu arac>l>¤>yla overeksprese olabilir veya li-
gandlar reseptörleri devaml> uyaracak flekilde eksprese olabilir. Mu-
tant ras proteinleri azalm>fl GTPaz aktivitesine veya GTPaz aktivite-
Bu de¤iflikliklerden herhangi birisi normal bir kontrol proteinini bir
onkoproteine dönüfltürebilir ve böylece neoplastik dönüflüme yol
açabilir.
Steroid Hormonlar
özellikleri çinko içeren DNA ba¤lay>c> bölgedir. Sistein rezidülerinin
aras>ndaki boflluk çinko atomu ile koordinasyon kurmaya elveriflli bir
yap> oluflturarak DNA'ya ba¤lan>r. Bir protein DNA etkilefliminin
enerjisi temas yüzeyine ba¤l> oldu¤undan proteinlerin ço¤u DNA'ya
komplex fleklinde ba¤lan>r. Glukokortikoit alt ailesi homodiner fleklin-
de tiroit hormon reseptör alt ailesi yine homodimer fleklinde fakat da-
ha s>kl>kla efllik eden RXR ile birlikte heterodimer fleklinde DNA'ya
ba¤lan>r (fiekil 240-5).
Glukokortikoit reseptör alt ailesinin üyeleri s>kl>kla palindromik böl-
gelere tutunurken tiroit hormon reseptör ailesi s>kl>kla tekrarlayan
DNA sekanslar>na do¤rudan ba¤lan>r. DNA ba¤layan bölge ve DNA
tan>ma bölgelerindeki küçük de¤ifliklikler hormon etkisinin özgüllü-
¤ünü sa¤lar. Reseptör ba¤lanmas>n>n önemli bir belirleyicisi dimerik
reseptör ba¤layan iki yar>m bölge aras>ndaki aral>kt>r. Tek aral>kl>
tekrarlar (DR+1) RXR homodimer ba¤lanmas> ve dokuz-sis-retinoik
fosforilasyon bölgesi, YP ba¤lay>c> Grb2'nin SH.2 domaini taraf>ndan farkedilir ve
ras'>n yerleflti¤i membrana guanin de¤iflim faktörü olan SOS'un gelmesini tetik-
ler. Aktifleflmifl GTP ba¤l> ras, serin threonin kinaz, olan rafile temasa geçerek
kinaz aktivasyonlar> kaskad>n> bafllat>r.