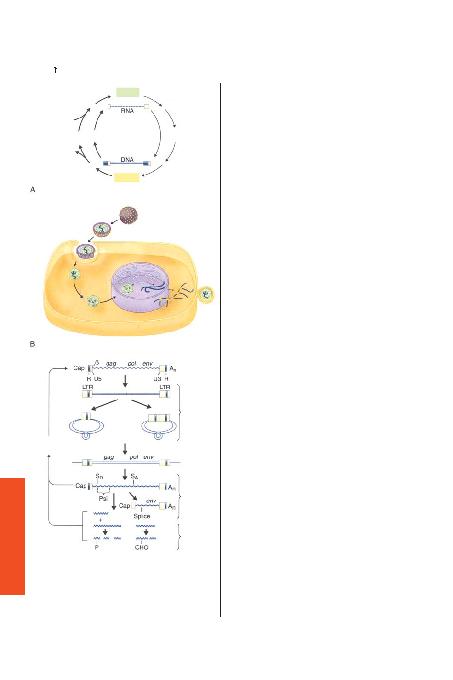
ile ba¤l> 70s kompleksi içinde birleflmifl ayn> iki RNA'dan oluflan dip-
loid bir yap>d>r. Bu genomik alt üniteler viral ürünleri oluflturan
mRNA ile ayn> kimyasal kutup içindeki artm>fl genomik viral RNA
parçalar>d>r. Ökaryotik mRNA'lardaki gibi genomik viral mRNA,
5'metil-G nükleotidi, 3'ucunda 100-200 nükleotidden oluflan poli-A
parças> ve birçok metillenmifl (A) rezidü kapsar. Konak hücre kaynak-
l> virion içine al>nan tRNA, viral genomik RNA'n>n 5'ucundaki primer
ba¤lanma yeriyle, 18 nükleotidden oluflan bir benzerli¤e sahiptir,
enfeksiyondan sonra viral replikasyonun ilk evrelerinde negatif DNA
sentezini bafllat>r.
ri kodlayan genleri (gag, pol ve env) içerir. Ek olarak HIV-1, virüslar>n
bu ailesine ait di¤er viral fonksiyonlar> kodlayan genleri de kapsar.
HIV-1 LTR sekans> viral genomun ekspresyonunu yönetir ve düzenler
(fiekil 409-5).
proteinler virüsün kor protein yap>s>na katk>da bulunur ve nükleik
asit ve lipid zar ba¤lanmas>na yard>mc> olur. Di¤er retrovirüslerde ol-
du¤u gibi HIV-1'in gag proteini viral matürasyon s>ras>nda parçala-
nan bir poliprotein öncülü olarak sentezlenir. Bu, spesifik bir viral pro-
teazla parçalanan viral kor yap>s>n>n bileflenlerinin 3 boyutlu olarak
toparlanmas>na yard>m eder böylece kor yap>s> olgun virionun özel-
leflmifl fonksiyonlar kazanmas>n> sa¤lar. Polimeraz gen ürünleri gag
proteinleri ile ayn> genomik RNA mesaj>nda oluflturulur, fakat ribozo-
mal proteinlerin flifti ile farkl> üst üste gelen bir nükleotid okuma dizi-
si oluflur. Pol geni büyük öncül polipeptidden parçalanan üç proteini
kodlar. Bu genler NH2-proteaz (p13)-revers transkriptaz (p66/p51)-in-
tegraz (p31)-COOH'I içerir. HIV-1 proteaz gag ve pol öncül polipep-
tidlerini fonksiyonel olarak aktif proteinlere parçalayarak virüs biyo-
lojisinde kritik bir rol oynar. Revers transkriptaz magnezyuma ihtiyaç
duyan ve viral RNA genomunun replikasyonundan sorumlu olan bir
RNA ba¤>ml> DNA polimerazd>r.
turan glikolize polipeptid öncül olan gp160'> kodlar. Son y>llarda
HIV'>n zarf>n>n yap>s> ve görevleri hakk>nda çok daha fazla fley ö¤re-
nilmifltir. Gp120/gp41 heterodimeri virüs yüzeyinde trimerik bir di-
ken halinde bulunur, iki ayr> molekül olarak görev yapmaktad>r, fakat
önemli görevleri virüsün ba¤lanmas> ve hücre içine giriflinin sa¤lan-
mas> ve virüs spesifik antikorlar ile nötralizasyondan kurtulunmas>-
d>r. Birinci görev gp120'nin s>rayla CD4 ve gp41 taraf>ndan gerçeklefl-
tirilen füzyon olay>n> tetikleyen kemokin reseptörüne ba¤lanmas> ile
yerine getirilir.Sonraki fonksiyon viral zarf yüzeyine maruz kalan kar-
bonhidrat bilefliminin gizlenmesi, maruz kalan viral zarf epitoplar>n>n
mutasyonu ve korunmufl zarf reseptör yüzeylerinin antikor ba¤lan-
mas>na karfl> konformasyonel veya enerjik bariyerler ile gizlenmesin-
den kaynaklan>r.
vpu genleridir. Bu genler pol ve env genleri aras>nda bulunur, nef ge-
ni env'in 3'taraf>ndad>r, ve viral LTR'nin U3 bölgesine uzan>r; ve tat ve
rev genleri virüsün merkezinde ve 3'ucunda yer alan iki parçal> kod-
lama ekzonu fleklindedir. Tat geni HIV-1 replikasyonu için gerekli
olan 14 kD `luk proteini kodlar. Bu protein HIV-1 ekspresyonunu
transkripsiyonel ve posttranskripsiyonel evrede artt>r>r. Tat'a ba¤l>
HIV-1 ekspresyonu art>fl>n>n hedef sekans> LTR'nin iflleme cevap ve-
ren bölgesidir (TAR). LTR TAR'> ba¤lamak ve aktiflefltirmek için hüc-
resel faktörlerle etkileflime girer (fiekil 409-5). Rev geni efllenmemifl ve-
ya tek olarak efllenmifl viral mRNA'n>n nükleustan sitoplazmaya ta-
fl>nmas> için gerekli 20-kD'lik bir proteini kodlar. Rev geni yoklu¤un-
da gag ve env proteinleri transkriptleri bölünerek parçalan>r ve böyle-
ce gag ve env proteinleri yap>lamaz. Vif geni antiretroviral selüler en-
zim olan apoB mRNA düzenleyici enzim, katalitik polipeptid benzeri
3G (APOBEC3G)'yi bask>layan 23 kD'luk bir protein ürününü kodlar
alt ünit
proteinler
ifllenmesi
transkriptaz