mas> ve vitamin D yan>t>n> yönlendirir, DR+4 tiroit hormon reseptö-
rü-RXR ba¤lanmas> ve tiroit hormon yan>t>n>, DR+5 ise retinoik asit
reseptör-RXR ba¤lanmas> ve atra yan>t>n> yönlendirir. Yar>m bölgeler
aras>ndaki aral>k Glukokortikoit reseptör s>n>f>n>n homodimerlerinin
ba¤lanmas> için gereklidir. Özgüllük mutlak de¤ildir fakat niceliksel-
dir. örne¤in progesteron reseptörleri Glukokortikoit yan>t birimine
ba¤lan>r, retinoik asit reseptörleri tiroit hormon reseptör yan>t birimi-
ne ba¤lan>r. Hormona özel yan>tlar>n oluflmas> için yeterli özgüllük
vard>r fakat Glukokortikoit regülasyonlu genlerin Ligand ile aktiflefl-
mifl reseptör indüksiyonunda oldu¤u gibi çak>flan fonksiyonlar orta-
ya ç>kabilir.
da bulunurken, kortizol ba¤lanmas> allosterik bir de¤ifliklikle disosi-
yasyona yol açarak ligand ba¤lanm>fl reseptörün DNA'ya tutunmas>-
na zemin oluflturur. Tiroit hormon ve retinoik asit reseptörleri ise di-
¤er proteinlerle komplex oluflturmay>p do¤rudan DNA'ya ba¤>ml>
halde bulunur. Hormon ba¤lanmas>yla tetiklenen allosterik de¤ifliklik
bu durumda reseptörü di¤er reseptörün di¤er transkripsiyon meka-
nizmas> ile etkileflime geçmesine yol açar. T
mas>n> tetikler. Tiroit hormon reseptörü ve aktivatör, CREB ba¤layan
proteinler gibi (CBP) baz> baflka proteinlerle etkileflerek multiple trans-
kripsiyon faktörleri (CREB, STAT gibi) ile tiroit hormon reseptör aile-
si aras>nda bilginin integrasyonunu sa¤lar.
asitleri, peroksizon proliferasyonuna yol açan metabolitler ve henüz
ligandlar> tan>mlanmam>fl öksüz reseptör için pek çok reseptör. Yeni
ligandlar keflfedildikçe baz> öksüz reseptörler dönüfltürülmektedir.
Örne¤in RXR ile heterodimer oluflturan reseptörler kolesterol, safra
asidi ve ksenobiotik metabolizmas>n> regüle etmektedir. Karaci¤erde
kolesterol-safra asidi metabolizmas> kolesterol katabolizmas>n>n ma-
jor yolu olup RXR reseptörlerine etki eden oxisteroitler taraf>ndan
uyar>l>r. (fiekil 240-6). LXR-
rü FXR bu aktivasyonu engeller böylece kolesterol katabolizmas> ve
safra asidi yap>m> aras>nda bir geri besleme oluflur. Bu reseptör ayn>
zamanda safra asidi geri al>m>n> kolaylaflt>ran proteinleri regüle
eder.
CYP2B gen transkripsiyonu üzerinden fenobarbital benzeri uyar>c>la-
ra yan>t> oluflturur. SXR/PXR ise ksenobiotikleri alg>lar ve CYP3A gen
ekspresyonunu uyararak ilaçlar>n ve toksik litokolik safra tuzlar>n>n
lerden toksik bileflenleri uzaklaflt>rmak için ABC tafl>y>c>lar>n>n eks-
presyonunu tetikler.
h>z>m> kontrol etmek üzere transkripsiyon bafllang>c>na göre farkl> po-
zisyonlardan etki gösterirler. Promoter genler ökaryoktik RNA poli-
meras II enziminin mRNA transkripsiyonunu bafllatt>¤> noktan>n üst
bölgelerinde yerlefliktir. En iyi tan>mlanm>fl promoter transkripsiyon
faktör II-D (TF-IID) proteinini ba¤layan bir TATA kutusu içerir ki bu,
30 baz çifti afla¤>daki RNA polimeras II taraf>ndan yönlendirilir. Yedi-
li proteinler (TATA ile iliflkili faktörler TAF) transkripsiyon kontrol
proteinleri ile etkileflime geçecek moleküler bir yüzey oluflturacak fle-
kilde TF II-D ile birliktedir. Di¤er promoter bileflenler bir bazal baflla-
t>c> ve guanin-sitozinden zengin bölgelerdir ki bunlar çoklu transkrip-
siyonun bafllad>¤> bölgelerdir. Transkripsiyon h>z> art>r>larak gen eks-
presyonu tetiklenir. Transkripsiyon h>z>n> artt>ran mekanizmalar; çok-
lu zay>f protein etkileflimlerinin toplam> ve DNA ile etkileflimi de¤iflti-
ren histon asetillenmeleridir.
Aktifleflmifl kortizol reseptörleri ACTH öncülünün kodlayan geni bas-
k>lar, aktifleflmifl tiroit hormon reseptörleri TSH-
pek çok mekanizma ile inhibe edebilirler örne¤in; histonlar>n DNA ile
etkileflimlerini artt>racak flekilde de-asetillenmesi gibi.
den oldu¤u gibi DNA'n>n özgül bölgelerine ba¤lan>rlar. Hücre yüze-
yinde oluflan hormonal sinyallere cevaben bu proteinler de¤iflebilir.
Böyle de¤ifliklikler hücre yüzeyine etki eden hormonlar>n oluflturdu¤u
gen transkripsiyon de¤iflikliklerine karfl>l>k gelir.
madan çekirde¤e yer de¤ifltirmesi. Siklik AMP ile kontrol edilen gen-
ler özgül çekirdek transkripsiyon regülatörlerini ba¤layan (CREB) se-
kanslar içerir. fosforilasyonu takiben aktivite s>ras>ndan de¤ifliklik ser-
gileyen CREB, hücre yüzeyinden etki gösteren adanilat siklaz>, aktive
eden peptit yap>l> hormonlar>n oluflturdu¤u gen uyar>s>n>n son arac>s>
olarak gereklidir. STAT ve iliflkili proteinler, tirozin bileflenleri üzerin-
den fosforlan>r ve fosforland>ktan sonra özgül genlerin transkripsiyo-
nunu aktive etmek üzere çekirde¤e girer. Bu zincirleme etkiler, mRNA
transkripsiyonunu de¤ifltirir, hücre fonksiyonu ve organ fizyolojisine
ait de¤ifliklikleri gerçeklefltirir.
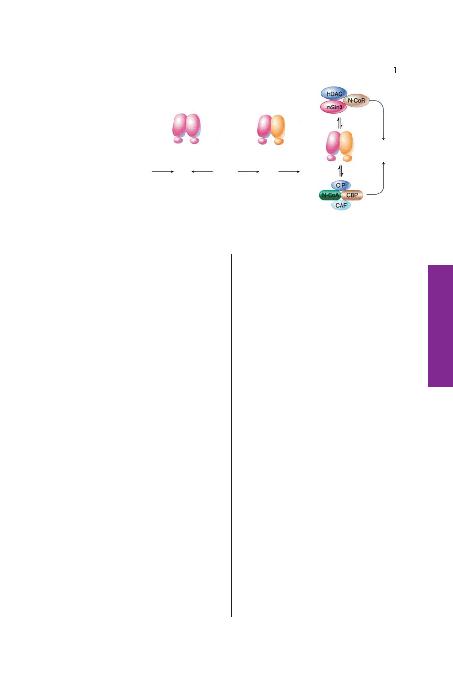
ailesi üyeleri polindramik DNA bölgelerine
ba¤lan>r. Tiroid hormon reseptör ailesi üye-
leri ise retinoid X reseptörleri ile heterodimer
oluflturup do¤rudan tekrarlanan DNA bölge-
lerine ba¤lan>r. Sa¤, hormon ba¤lanmas>n>
takiben, repressör kompleksi çözülür ve ak-
tivatör kompleks nükleer reseptörlere ba¤la-
n>r. repressör kompleksler histon deasetilaz
(HDAC) aktivatör kompleksler ise histon
asetilaz (CAF) içerir. Koaktivatör ve korep-
ressar kompleksler çoklu proteinler içerir,
yaln>zca bir k>sm> burada gösterilmifltir.