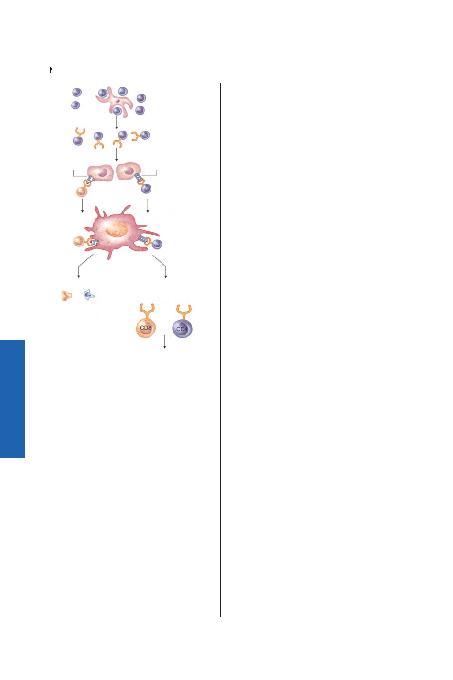
CD80/CD86 ligandlar>na ba¤lanan CD28 molekülüdür. CD28 arac>l>
sinyaller, baflta IL-2 salg>lanmas> olmak üzere, yan>t veren T hücreler
üzerinde bir çok aktivasyon molekülünün ekspresyonu için zorunlu-
dur. TCR'dan gelen bir çok sinyal bir çok genin aktivasyonu ve T hüc-
renin hücre döngüsüne girmesine neden olur. Sinyaller bir sitoplaz-
mik olaylar kaskad> ile iletilir. TCR ve onunla iliflkili CD3 molekülü-
nün çapraz ba¤lanmas> (cross-link) fosfotirozin kinaz>n çal>flt>r>lmas>
ve aktivasyonu ve TCR'ün moleküler içeri¤inin ve farkl> adaptör mo-
leküllerin fosforilasyonu ile sonuçlan>r. TCR arac>l>¤> ile iletilen sinyal-
ler, gen ekspresyonunu düzenleyen transkripsiyon faktörlerinin akti-
vasyonu ile sonuçlanan MAP-kinaz, protein-kinaz C ve kalsinörin yo-
lu; fosfotidil inozitol ürünlerinin oluflturulmas> ve hücre içi kalsiyu-
mun artt>r>lmas> gibi bir çok biyokimyasal yolu aktive eder.
rasyon evresi, antijen sunan hücrenin do¤as>. T hücre yan>tlar>n>n bafl-
lamas>nda antijen sunan hücreler önemli rol oynarlar. Kostimülatör
sinyaller oluflturan aksesuvar moleküllerin ekspresyonunu artt>r>rlar.
Dendritik hücreler üzerinde MHC/peptid komplekslerinin özellikle
yo¤un olmas>, bu hücrelerin naive T hücreleri aktive etmesini sa¤lar.
Bunun aksine, bellek ve efektör hücrelerin aktivasyon efli¤i düflüktür
ve periferik doku hücreleri taraf>ndan sunulan antijene tepki verebilir.
Klonal proliferasyonun boyutu son derece etkileyicidir. Antijen spesi-
fik CD8
2 a¤>rl>kl> otokrin mekanizmalarca yürütülen T hücreler, h>zl> büyüme
evresinde, neredeyse hiç bir efektör fonksiyonu olmayan naive T hüc-
relerden efektör T hücrelere farkl>lafl>r. Efektör hücreye geçifl, temel bir
fonksiyonel kayma ile oluflur. fiunu belirtmek gerekir ki; öncelikle,
efektör T hücrelerin aktivasyon efli¤i düflüktür; kostimülasyon gerek-
tirmezler ve profesyonel antijen sunan hücreleri içermeyen dokular>
da tarayabilirler, ikinci olarak, periferik dokulara ulaflabilmek için Ke-
mokin reseptörleri ve adezyon moleküllerinin ekspresyonunu de¤iflti-
rebilirler. En sonunda da efektör fonksiyon kazan>rlar, CD8
CD4
çok sitokin salg>larlar. CD8
(TH1) ad> verilen CD4
olan IL-4, IL-5 ve IL-13 üretirler. Hangi farkl>laflma yolunun seçilece¤i
T hücrere aktivasyonunun erken evrelerinde belirlenir ve mikro çevre-
de bulunan do¤al immün sistem hücrelerince yap>lan sitokinler, ko-
stimülator sinyallerin do¤as> ve TCR-MHC/antijen iliflkisinin avidite-
si gibi birçok faktöre dayan>r.
s>n>n oldu¤u bölgeye toplan>r; temas alan>nda özelleflmifl litik granül-
ler birikir. Por oluflturucu bir protein olan perforin, litik granüllerden
sal>narak hedef hücre membran>na sald>r>r. Proteazlar (granzimler)
enzim kaskadlar>n> aktive ederek apoptotik olay> bafllatmak için hedef
hücre içine verilirler. CD8
den olabilir ve buradaki mekanizma farkl>d>r. Aktive olmufl CD4
apoptotik kaskad> bafllat>r. CD4
hücre taraf>ndan tan>nmas> sonucunda ortaya ç>kar. T hücrelerin hedef
hücrede toplanmas>, sitokinlerin sal>n>m> ve stimulatör hücre yüzey
moleküllerinin ekspresyonu izler. Th1 ve Th2 lenfositler, B hücrelere
yard>m etmek için ise, B hücre yüzeyindeki CD40'> stimüle etme ama-
c>yla CD40 ligand (CD154) eksprese ederler. TH1 hücreler, B hücreleri
IgGl ve IgG3 s>n>f>ndan opsonizan antikorlar yap>m> için aktive eder;
TH2 hücreler ise üzerinde IgM tafl>yan B hücre yan>tlar>n>n bafllat>lma-
s>ndan ve tüm di¤er s>n>flardan immünglobülinlerin özellikle IgE ya-
p>m>n>n desteklenmesinden sorumludur.
kompartman>nda yer s>n>rl>d>r. Yer ve kaynaklar için yar>fl> ve hayat
boyu antijen maruziyeti ile T hücre çeflitlili¤inin bozulmas>n> engelle-
mek için, kazan>lm>fl immün sistem baz> karfl> mekanizmalar uygular.
Aktivasyon olay> s>ras>nda, T hücre yüzey moleküllerinden biri olan
CTLA-4 (CD152) ve antijen sunan hücreler üzerindeki CD80/CD86
aras>ndaki iliflkiden güçlü bir negatif sinyal oluflur. Ayr>ca, CD4
CD4
kar. Bu mekanizmalar T hücre antijen yan>tlar>n>n erken evrelerinde
k>s>tlamalar ortaya ç>kar>r. Afla¤>da anlat>lan di¤er mekanizmalar ise
antijenin ortadan kald>r>lmas> baflar>ld>¤>nda genifllemifl antijen-spesi-
sa geldiklerinde T hücre reseptör genleri için tekrar düzenlenme bafllar. Kortikal
epitelyal hücrelerdeki kendi MHC kompleksine bafllanan immatür T hücreleri ya-
flamlar>n> sürdürmek için sinyaller al>rlar (pozitif seçim). Kortikomedüller bileflke-
de makrofaj ve dendritik hücreler taraf>ndan yaflamlar>n> sürdüren bu T hücrele-
rine kendi antijenleri sunulur. Kendi antijenlerine kuvvetli yan>t veren T hücreler
apopitozis yoluyla ortadan kald>r>l>rlar (negatif seçim). Kendi antijenlerine toleran
yabanc> antijenleri ise tan>yan T hücreler perifere sal>n>rlar.
hücre
kendi antijeni
self tan>ma